Шпаргалка по расчету налога на прибыль и авансовых платежей за I квартал
Предприятия уплачивают авансовые платежи по налогу на прибыль ежемесячно либо ежеквартально. Периодичность уплаты налога...
Включают следующие ныне существующие отделы: моховидные (Bryophyta ), плауновидные (Lycopodiophyta ), псилотовидные (Psilotophyta ), хвощевидные (Equisetophyta ), папоротниковидные (Polypodiophyta ).
Споровые растения появились в конце силурийского периода, более 400 млн. лет тому назад. Первые представители споровых были небольших размеров и имели простое строение, но уже у примитивных растений наблюдалась дифференциация на элементарные органы. Усовершенствованию органов соответствовало усложнение внутреннего строения и онтогенеза. В жизненном цикле происходит чередование полового и бесполого способов размножения и связанное с этим чередование поколений. Бесполое поколение представлено диплоидным спорофитом , половое – гаплоидным гаметофитом .
На спорофите образуются спорангии, внутри которых в результате мейотического деления образуются гаплоидные споры. Это мелкие, одноклеточные образования, лишенные жгутиков. Растения, у которых все споры одинаковые, называются равноспоровыми. У более высокоорганизованных групп споры двух типов: микроспоры (формируются в микроспорангиях), мегаспоры (формируются в мегаспорангиях). Это разноспоровые растения. При прорастании споры образуется гаметофит.
Полный жизненный цикл (от зиготы до зиготы) состоит из гаметофита (период от споры до зиготы) и спорофита (период от зиготы до образования спор). У плаунов, хвощей и папоротников эти фазы представляют собой как бы отдельные физиологически самостоятельные организмы. У мхов гаметофит является самостоятельной фазой жизненного цикла, а спорофит сведен до его своеобразного органа – спорогона (спорофит живет на гаметофите).
На гаметофите развиваются органы полового размножения: архегонии и антеридии . В архегониях , похожих на колбу, образуются яйцеклетки, а в мешковидных антеридиях - сперматозоиды. У равноспоровых растений гаметофиты обоеполые, у разноспоровых – однополые. Оплодотворение происходит лишь при наличии воды. При слиянии гамет образуется новая клетка – зигота с двойным набором хромосом (2n).
Отдел Моховидные – Bryophyta
Насчитывает до 27000 видов. Моховидные имеют тело либо в виде слоевища, либо расчленены на стебель и листья. Настоящих корней у них нет, их заменяют ризоиды. Проводящие ткани появляются лишь у высокоразвитых мхов. Частично обособлены ассимиляционная и механическая ткани.
В жизненном цикле доминирует гаметофит. Спорофит самостоятельно не существует, развивается и всегда находится на гаметофите, получая от него воду и питательные вещества. Спорофит представляет собой коробочку, где развивается спорангий, на ножке, связывающей ее с гаметофитом.
Размножаются мхи спорами, могут размножаться и вегетативным путем – отдельными участками тела или особыми выводковыми почками.
Отдел делится на три класса : Антоцеротовые (100 видов, шесть родов талломных растений), Печеночные и Листостебельные мхи.
Класс Печеночные мхи (Hepaticopsida )
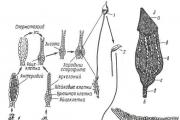
Класс насчитывает около 8500 видов. Это, в основном, слоевищные мхи, хотя есть виды, имеющие стебель и листья. Широко распространена маршанция обыкновенная (Marchantia polymorpha ) (рис 11. 1).
Рис. 11. 1. Цикл воспроизведения маршанции: 1 – таллом с мужскими подставками; 2 – таллом с женскими подставками; 3 – вертикальный разрез через мужскую подставку (в некоторых антеридиальных полостях находятся антеридии); 4 – антеридий в антеридиальной полости (н – ножка антеридия); 5 – двужгутиковый сперматозоид; 6 – вертикальный разрез через женскую подставку (а – архегоний).
Гаметофит имеет темно-зеленое слоевище (таллом), дихотомически разветвленное на широкие лопастные пластинки с дорсовентральной (спинно-брюшной) симметрией. Сверху и снизу слоевище покрыто эпидермой, внутри имеются ассимиляционная ткань и клетки, выполняющие проводящую и запасающую функции. К субстрату слоевище прикрепляется ризоидами . На верхней стороне слоевища в особых «корзиночках» образуются выводковые почки, служащие для вегетативного размножения.
Талломы раздельнополые, органы полового размножения развиваются на особых вертикальных ветвях-подставках.
Мужские гаметофиты имеют восьмилопастные подставки, на верхней стороне которых находятся антеридии . На женских гаметофитах подставки со звездчатыми дисками, на нижней стороне лучей звездочки расположены (шейкой вниз) архегонии. При наличии воды сперматозоиды перемещаются, попадают в архегоний и сливаются с яйцеклеткой.
После оплодотворения из зиготы развивается спорогон. Он имеет вид шаровидной коробочки на короткой ножке. Внутри коробочки, в результате мейоза, из спорогенной ткани, образуются споры. В благоприятных условиях споры прорастают, из них развивается протонема в виде небольшой нити, из верхушечной клетки которой развивается таллом маршанции.
Класс Листостебельные мхи (Bryopsida, или Musci ).
Листостебельные мхи распространены по всему земному шару, особенно в условиях холодного климата на сырых местах, в сосновых и еловых лесах, в тундре. На торфяных и моховых болотах часто образуют плотный ковер. Тело расчленено на стебель и листья, но настоящих корней нет, есть многоклеточные ризоиды. Класс состоит из трех подклассов: Бриевые, или Зеленые мхи; Сфагновые, или Белые мхи; Андреевые, или Черные мхи.
Андреевые мхи (три рода, 90 видов) распространены в холодных областях, внешне сходны с зелеными, по строению листьев и коробочки – со сфагновыми.
Подкласс Бриевые, или Зеленые мхи (Bryidae ). Насчитывает около 700 родов, объединяющих 14000 видов, широко распространенных повсюду, особенно в тундровой и лесной зонах Северного полушария.
Широко распространен кукушкин лен (Polytrichium commune ), образующий густые дерновины на сырых почвах в лесах, на болотах и лугах. Стебли до40 см высоты, неветвистые, с густыми жесткими и острыми листьями. От нижней части стебля отходят ризоиды.
Цикл развития кукушкина льна (рис. 11. 2).

Рис. 11. 2. Кукушкин лен: А – цикл развития мха; Б – коробочка: 1 – с колпачком, 2 – без колпачка, 3 – в разрезе (а –крышечка, б – урночка, в – спорангий, г – апофиза, д – ножка); В – поперечный разрез листа с ассимиляторами; Г – поперечный разрез стебля (ф – флоэма, крв – крахмалоносное влагалище, кор – кора, э – эпидерма, лс – листовые следы).
Гаметофиты кукушкина льна раздельнополые. Рано весной на верхушке мужских особей развиваются антеридии, на верхушках женских – архегонии.
Весной во время дождя или после росы сперматозоиды выходят из антеридия и проникают в архегоний, где сливаются с яйцеклеткой. Из зиготы здесь же, на верхушке женского гаметофита, вырастает спорофит (спорогон), имеющий вид коробочки на длинной ножке. Коробочка покрыта волосистым колпачком (калиптра) (остатком архегония). В коробочке – спорангий, где после мейоза образуются споры. Спора – небольшая клетка с двумя оболочками. На верхушке коробочки, по ее краю, расположены зубцы (перистом), которые, в зависимости от влажности воздуха, загибаются внутрь коробочки или отгибаются наружу, чем способствуют рассеиванию спор. Споры разносятся ветром и в благоприятных условиях прорастают, образуя протонему. Через некоторое время на протонеме образуются почки, из которых формируются листостебельные побеги. Эти побеги вместе с протонемой – гаплоидное поколение – гаметофит. Коробочка на ножке – диплоидное поколение – спорофит.
Подкласс Сфагновые, или Белые мхи (Sphagnidae)
К сфагновым мхам относятся свыше 300 видов единственного рода сфагнум (Sphagnum ) (рис. 11. 3).

Рис 11. 3. Сфагнум : 1 – внешний вид; 2 – верхушка ветви со спорогоном; 3 – спорогон (ш – остаток шейки архегония, кр – крышечка, сп – спорангий, кол – колонка, н – ножка спорогона, лн – ложная ножка); 4 – часть веточного листа (хлк – хлорофиллоносные клетки, вк – водоносные клетки, п – поры); 5 – поперечный разрез листа.
Ветвистые стебли сфагнума усеяны мелкими листьями. На верхушке главной оси боковые веточки образуют розетку почковидной формы. Особенностью сфагновых мхов является непрерывное нарастание стебля верхушкой и отмирание нижней части. Ризоиды отсутствуют, и поглощение воды с минеральными веществами происходит стеблями. Листья этих мхов состоят из двух типов клеток: 1) живых ассимилирующих, длинных и узких, хлорофиллоносных; 2) гиалиновых – мертвых, лишенных протопласта. Гиалиновые клетки легко заполняются водой и долго ее сохраняют. Благодаря такому строению, сфагновые мхи могут накапливать воды в 37 раз больше своей сухой массы. Разрастаясь плотными дернинами, сфагновые мхи способствуют заболачиванию почв. На болотах наслаивание отмерших частей мха приводит к образованию торфяников. Путем сухой перегонки из торфа получают воск, парафин, фенолы, аммиак; путем гидролиза – спирт. Торфяные плиты являются хорошим теплоизоляционным материалом. Сфагновые мхи обладают бактерицидными свойствами.
Отдел Плауновидные – Lycopodiophyta
Появление плауновидных связывают с силурийским периодом палеозойской эры. В настоящее время отдел представлен травянистыми растениями со стелющимися, дихотомически ветвящимися стеблями и корнями, а также спирально расположенными чешуйчатыми листьями. Листья возникли как выросты на стебле, и их называют микрофиллы . У плаунов есть флоэма, ксилема, перицикл.
Выделяют два современных класса: равноспоровые Плауновые и разноспоровые Полушниковые.
Класс Плауновые (Lycopodiopsida)
Из всего класса до настоящего времени сохранилось четыре рода.
Род плаун (Lycopodium). К этому роду относят многочисленные (около 200 видов) многолетние вечнозеленые травы, распространенные от арктических областей до тропиков. Так плаун булавовидный (L. clavatum) встречается в травяном покрове хвойных лесов на достаточно влажных, но бедных гумусом почвах. В сырых хвойных лесах широко распространен плаун годичный (L. annotinum ) (рис. 11. 4).

Рис. 11. 4. Плаун булавовидный.
Родбаранец (Huperzia). Представитель рода - баранец обыкновенный (H. selago) распространен в тундровой, лесотундровой и северной лесной зонах и произрастает в южно-таежных еловых лесах и ольшаниках, а также в мшистых лесах и на альпийских лугах.
Роддифазиаструм (Diphasiastrum ). Представитель рода дифазиаструм сплюснутый (D. complanatum) растет на сухих песчаных почвах в сосновых лесах.
Цикл развития на примере плауна булавовидного (рис. 11. 5).

Рис. 11. 5. Цикл развития плауна булавовидного :1 – спорофит; 2 – спорофилл со спорангием; 3 – спора; 4 – гаметофит с антеридиями и архегониями; 5 – развивающийся на гаметофите из зародыша молодой спорофит.
Стелющиеся побеги плауна булавовидного достигают до 25 см в высоту и более 3 м в длину. Стебли покрыты спирально расположенными ланцетно-линейными мелкими листьями. В конце лета на боковых побегах образуется обычно два спороносных колоска. Каждый колосок состоит из оси и мелких тонких спорофиллов – видоизмененных листьев, в основании которых расположены спорангии почковидной формы.
В спорангиях после редукционного деления клеток спорогенной ткани образуются одинаковой величины, одетые толстой желтой оболочкой гаплоидные споры. Они прорастают после периода покоя через 3-8 лет в обоеполые заростки, представляющие собой половое поколение и живущие сапротрофно в почве, в виде клубенька. От нижней поверхности отходят ризоиды. Через них в заросток врастают гифы гриба, образуя микоризу . В симбиозе с грибом, который обеспечивает питание, живет заросток, лишенный хлорофилла и неспособный к фотосинтезу. Заростки многолетние, развиваются очень медленно, лишь через 6-15 лет на них образуются архегонии и антеридии. Оплодотворение совершается при наличии воды. После оплодотворения яйцеклетки двужгутиковым сперматозоидом образуется зигота, которая без периода покоя прорастает в зародыш, развивающийся во взрослое растение.
В официнальной медицине споры плауновприменялись в качестве детской присыпки и обсыпки для пилюль. Побеги баранца обыкновенного используют для лечения больных, страдающих хроническим алкоголизмом.
Класс Полушниковые (Isoetopsida)
Селагинелла (Selaginella ) среди современных родов насчитывает самое большое (около 700) число видов.
Это нежное многолетнее травянистое растение, требующее высокой влажности. Селагинеллы в отличие от плаунов характеризуются разноспоровостью. В спороносных колосках образуется два вида спор – четыре мегаспоры в мегаспорангиях и многочисленные микроспоры в микроспорангиях. Из микроспоры образуется мужской гаметофит, состоящий из одной ризоидальной клетки и антеридия со сперматозоидами. Мегаспора развивается в женский гаметофит, не покидающий ее оболочки и состоящий из мелкоклеточной ткани, в которую погружены архегонии. После оплодотворения из яйцеклетки развивается зародыш, и затем новый спорофит.
Отдел Хвощевидные – Equisetophyta
Появились хвощи в верхнем девоне, достигли наибольшего разнообразия в карбоне, когда древесный ярус заболоченных тропических лесов в значительной степени состоял из древовидных хвощевидных, вымерших к началу мезозоя. Современные хвощевидные появились на Земле с мелового периода.
До настоящего времени сохранился лишь один род – хвощ (Equisetum), представленный 30-35 видами, распространенными на всех континентах.
У всех видов хвоща стебли имеют членистое строение с выраженным чередованием узлов и междоузлий. Листья редуцированы до чешуй и располагаются мутовками в узлах. Здесь же образуются и боковые ветви. Ассимилирующую функцию выполняют зеленые стебли, поверхность которых увеличивается ребристостью, стенки клеток эпидермы пропитаны кремнеземом. Подземная часть представлена сильно развитым корневищем, в узлах которого формируются придаточные корни. У хвоща полевого (Equisetum arvense) боковые ветви корневища служат местом отложения запасных веществ, а также органами вегетативного размножения (рис. 11. 6).

Рис. 11. 6. Хвощ полевой : а, б – вегетативный и спороносный побеги спорофита; в – спорангиофор со спорангиями; г, д – споры; е – мужской гаметофит с антеридиями; ж – сперматозоид; з – обоеполый гаметофит; и - архегонии.
Весной на обычных или специальных спороносных стеблях образуются колоски, состоящие из оси, которая несет особые структуры, имеющие вид шестигранных щитков (спорангиофоры ). Последние несут 6-8 спорангиев. Внутри спорангиев образуются споры, одетые толстой оболочкой, снабженной гигроскопичными лентовидными выростами – элатерами. Благодаря элатерам споры сцепляются вместе в комки, хлопья. Групповое распространение спор способствует тому, что при их прорастании разнополые заростки оказываются рядом, и это облегчает оплодотворение.
Заростки имеют вид небольшой длинно-лопастной зеленой пластинки с ризоидами на нижней поверхности. Мужские заростки меньше женских и несут по краям лопастей антеридии с многожгутиковыми сперматозоидами. На женских заростках в средней части развиваются архегонии. Оплодотворение происходит при наличии воды. Из зиготы развивается зародыш нового растения – спорофита.
В настоящее время большой роли в образовании растительного покрова хвощи не имеют. В лесах, на избыточно увлажненной почве, широко распространен хвощ лесной (E. sylvaticum) с сильно ветвящимися, поникающими боковыми веточками. На лугах, паровых полях, в посевах встречается трудно искореняемый сорняк хвощ полевой (E. arvense). У этого хвоща рано весной появляются неветвящиеся побеги, несущие спороносные колоски. Позднее от корневища развиваются зеленые вегетативные побеги. В лесной зоне на песчаных почвах и в оврагах широко распространен хвощ зимующий (E. hyemale ).
Вегетативные побеги хвоща полевого (E. arvense) в официнальной медицине применяют: в качестве мочегонного средства при отеках на почве сердечной недостаточности; при заболеваниях мочевого пузыря и мочевыводящих путей; как кровоостанавливающее средство при маточных кровотечениях; при некоторых формах туберкулеза.
Отдел Папоротниковидные – Polypodiophyta
Возникли папоротники в девоне, когда древовидные папоротники вместе с ныне ископаемыми плаунами и хвощами господствовали в растительном покрове земли. Большинство их вымерло, оставшиеся дали начало мезозойским формам, которые были представлены очень широко. Папоротники намного превосходят по числу современных видов все остальные отделы высших споровых (около 25000).
У большинства живущих сейчас (исключая тропические) папоротников отсутствует наземный прямостоячий стебель, но есть подземный в виде корневища. От корневища отходят придаточные корни и крупные листья (вайи ), имеющие стеблевое происхождение и длительно нарастающие верхушкой. Молодые листья обычно свернуты «улиткой». Среди существующих в настоящее время папоротников имеются как равноспоровые, так и разноспоровые.
В лесах Австралии, Южной Америки, Азии произрастают древовидные представители с колонновидными, неветвящимися, до 20 метров высоты стволами. В средней полосе нашей страны папоротники – это многолетние корневищные травы. Многие папоротники, как и мхи, являются индикаторами почв и типов леса. В светлых лесах, на песчаных или сухих подзолистых почвах распространен орляк обыкновенный (Pteridium aquilinum); на влажных богатых почвах кочедыжники (Athyrium) и крупныелесныещитовники (Dryopteris) (рис. 11. 7).

Рис. 11. 7. Щитовник мужской: А – спорофит: а – общий вид; б – сорусы на нижней стороне вайи; в – разрез соруса (1 – индузий, 2 – планцента, 3 – спорангий); г – спорангий (4 – кольцо); Б – гаметофит: 5 – сперматозоиды; 6 – заросток с нижней стороны (т – таллом, р – ризоиды, арх - архегонии, ан – антеридии); 7 – выход сперматозоидов из антеридия; 8 – архегоний с яйцеклеткой.
Цикл развития равноспоровых папоротников
В середине лета на нижней стороне зеленых листьев (у некоторых на специальных спороносных листьях) в виде коричневых бородавочек появляются группы спорангиев (сорусы) . Сорусы у многих папоротников прикрыты сверху своеобразным покрывалом - индузием. Спорангии образуются на особом выросте листа (плаценте) и имеют чечевицеобразную форму, длинные ножки и многоклеточные стенки. В спорангиях хорошо выражено механическое кольцо, которое имеет вид узкой несмыкающейся полосы, опоясывающей спорангий. Когда кольцо подсыхает, стенки спорангия разрываются, и споры высыпаются.
Споры, образовавшиеся в спорангиях, одноклеточны и имеют толстую оболочку. При созревании они разносятся током воздуха и в благоприятных условиях прорастают, образуя сердцевидную зеленую многоклеточную пластинку (заросток), прикрепляющуюся к почве ризоидами. Заросток представляет собой половое поколение папоротников (гаметофит). На нижней стороне заростка образуются антеридии (со сперматозоидами) и архегонии (с яйцеклетками). При наличии воды сперматозоиды проникают в архегонии и оплодотворяют яйцеклетки. Из зиготы развивается зародыш, имеющий все основные органы (корень, стебель, лист и особый орган – ножку, прикрепляющую его к заростку). Постепенно зародыш начинает существовать самостоятельно, а заросток отмирает.
У разноспоровых папоротников гаметофиты редуцированы до микроскопических размеров (особенно мужские).
Из корневищпапоротника мужского (Dryopteris filix-mas), получают густой экстракт, который является эффективным противоглистным средством (ленточные черви).
Определение 1
Высшие споровые растения - это растения, которые населяют наземную среду и размножаются спорами.
Высшие споровые растения - это новый этап в эволюционном развитии растений. Высшим растениям, в отличие от низших, присуще расчленение тела на вегетативные органы: корень, листья и стебель . Вегетативные органы построены из разнообразных тканей.
Все высшие споровые растения, как правило, обитатели суходола, но среди них есть и обитатели водоёмов.
К высшим споровым растениям относятся все наземные листостеблевые растения, которые размножаются спорами. Это представители отделов:
Высшие растения появились в далёкой древности. Возможными их предками были бурые или зелёные водоросли, которые достигли высокой дифференциации тела и органов полового размножения.
Водная среда заселена в основном низшими растениями, тогда как на суше господствующими есть высшие, у которых с выходом на сушу образовался ряд характерных приспособлений к новой среде.
Наличие разных типов тканей
Внешне растения покрыты тканью, защищающей их от неблагоприятных условий. Процесс фотосинтеза обеспечивается хорошо развитой хлорофиллоносной тканью. Благодаря наличию проводящих тканей происходит обмен веществ между подземными и надземными органами. Кроме того, хорошо развиты механические (опорные) и запасающие ткани.
Расчленение тела на органы
Прежде всего у высших растений сформировались специальные органы поглощения минеральных веществ из субстрата - ризоиды и корневые волоски. Поскольку общей биологической чертой всех высших растений является автотрофное питание, у них образовался фотосинтезирующий орган - листок. Стебель и корень сформировались для связи двух важных конечных аппаратов - корневого волоска и зелёной клетки листка, а так же для обеспечения стойкости растения в воздушной среде и почве.
Органы полового размножения всегда многоклеточные
Бывают двух типов: мужские (антеридии) и женские (архегонии).
Онтогенез начинается из зародыша
Зародыш развивается из зиготы - клетки, которая образуется в результате слияния гамет.
Наличие правильного чередования полового и бесполого поколений
Половое поколение называется гаметофит, бесполое - спорофит.
Преобладание в цикле развития спорофита (за исключение мохообразных)
Прогрессивное развитие спорофита вызвано высокой приспособленностью к наземным условиям и великим объёмом генетической информации, свойственной диплоидному поколению.
Определение 2
Жизненным циклом называется черёд фаз в развитии растения, по завершении которых растительный организм достигает зрелости и становится способным давать начало будущим поколениям (половому - гаметофиту и бесполому - спорофиту).
Жизненный цикл высших споровых растений состоит из ритмического чередования двух поколений: бесполого (спорофита) и полового (гаметофита).
На спорофите образуются спорангии - органы бесполого размножения, в которых формируются споры. Из них развиваются гаметофиты, то есть особи полового поколения, которые могут быть одно- и двуполыми.
На гаметофите образуются антеридии - мужские половые органы и архегонии - женские. В антеридиях формируются подвижные сперматозоиды, а в архегониях - неподвижные яйцеклетки.
Оплодотворение возможно только при наличии капельножидкой воды. Вода необходима для передвижения сперматозоида к яйцеклетке. Из оплодотворённой яйцеклетки развивается зародыш, который растёт и превращается на особь бесполого поколения - спорофит.
Следует помнить, что из зиготы развивается лишь спорофит, а из споры - лишь гаметофит.
Замечание 1
В цикле развития большинства споровых растений (за исключением мохообразных) преобладает спорофит, который значительно лучше приспособлен к жизни в сложных условиях наземной среды. То есть, для эволюции высших споровых растений (кроме мхов) характерна тенденция к преобладанию и усовершенствованию спорофита при одновременной редукции гаметофита.
В лесу многие из этих растений, часто мелких, обращают на себя внимание большей формой, и ни одно - яркими цветами, что дало повод назвать их нецветковые. Второе название, споровые, они получили за свои органы размножения - споры - одноклеточные микроскопические образования, которые в начале прорастания дают заросток, встреча мужских и женских клеток, и только потом появляется само зеленое растение.
Мир споровых растений включает в себя несколько классов, начиная от высших: папоротники, хвощи, плауны, мхи, и заканчивая низшими - лишайниками. Из всего обилия споровых растений тайги самые крупные - папоротники. Из них обычны: кочедыжник женский, голокучник Линнея, страусник.
С наступлением тепла, когда все растения распускают свои листья, оживают и папоротники, при этом проявляют свою особенность, выпуская вначале из земли стебель в виде зеленых спиралей, чем отличаются от семенных, у которых росток устремляется прямо вверх.
Через несколько недель, когда в листьях накопится достаточное количество энергии, наступает пара размножения, и появляются особые образования сорусы, в которых развиваются споры. Они располагаются на нижней стороне листьев и сверху не видны. Чтобы увидеть их, нужно листья повернуть, и тогда перед глазами они предстанут в виде коричневатых почек или бугорков.
Расположение сорусов возле жилок листа, форма их краев, количество - все это служит дополнительным отличительным признаком одного вида папоротника от другого.
Из крупных папоротников самый обычный - кочедыжник женский. Свое название кочедыжник он получил за крючочки, расположенные в основании листьев, напоминающих своей формой крупные крючки, с помощью которых деревенские жители плели лапти. Кочедыжник часто образует заросли из широких, сложно изрезанных, перистых, склоненных к земле листьев. Когда проходишь через такие заросли, то как будто на мгновение попадаешь в зеленый мир древнего каменноугольного периода - время господства споровых растений.
Другой высокий папоротник, страусник, встречается реже и предпочитает лога с ручьями. Листья у него тоже перистые, но в отличие от кочедыжника, поднимаются прямо вверх, почти до метра, и располагаются по кругу, образуя зеленый конус, похожий на стеклянный сосуд, который французы называют "фужер" . Еще одно отличие, заметное издалека: его сорусы собираются на отдельном листе, который своей формой напоминает перо страуса, правда, коричневого цвета.
Третий обычный папоротник - голокучник Линнея, который раньше называли щитовник из-за треугольной формы листьев, напоминающей щиты древних воинов. Он невысокий, от 15 до 20 см, если летом он как-то теряется среди зелени травы, то осенью становится далеко заметным, принимая беловатую окраску.
Следующий класс споровых растений - хвощи. Самый высокий и необычный по форме из них - зимующий. Заросли его напоминают зеленые трубки высотой до 60 см. Для них характерно, что они почти не меняют свой зеленый цвет, осенью среди желтой листвы они выглядят свежими, весной, пробиваясь из-под снега, более темными, но зелеными.
После окончательного схода снега, когда поверхность земли прогреется, зимующий хвощ оживает, признаком этого служит появление в местах соединения отдельных члеников стебля (членистость стеблей - характерный признак всех хвощей) - мелких острых листочков. Другой признак оживления - разрастание на кончике стебля коричневатой шишечки - спорангия.
В жизни леса зимующий хвощ имеет значение как кормовая трава для лося, которая рано выходит из-под снега. Кроме этого, зимующий хвощ находил применение в промышленности, и связано это было с особенностью строения его клеток. В них, как и у других представителей хвощовых, содержатся кристаллы твердого минерала кремния, но в особо концентрированном состоянии. Это преимущество и было использовано для шлифовки оптических стекол стеблями, когда не было специальных паст.
Другой хвощ, с особенностями которого пришлось познакомиться детям в далекие годы военного лихолетья, когда были затруднения с пищей - лесной. Для этого хвоща характерно, что при наступлении тепла зимующие корневища вначале выпускают голые стебли с беловатыми острыми шишечками, наполненными жирными спорами. Именно эти шишечки, под названием "пестики", и шли тогда в пищу.
В июне стебли лесного хвоща покрываются зелеными веточками-листьями, расположенными круговыми ярусами, от чего само растение приобретает оригинальный вид, а заросли его придают лесным полянам какой-то туманный цвет, как будто на траве лежит нежная вуаль из тонких нитей.
На лесной хвощ походит полевой, у которого тогда ценились корневые клубеньки. Их обычно собирали по береговым обрывчикам. На вкус они были сладковатые и цвет имели черный. От лесного хвоща полевой отличался тем, что его листья-веточки ветвились один раз, и все растение выглядело не таким густым.
Полевой хвощ обычно растет в поймах рек, где образует настолько обширные поля по сырым ложбинам, что они получили название хвощевники, которые в пору заготовки кормов выкашивали для получения особого сена. Это сено, обладая положительными для желудка скота свойствами, имело отрицательное значение для зубов: при длительном потреблении быстро стирало эмаль из-за наличия кремния в клеточках.
Все хвощи предпочитают сырые места, а топяной - прямо воду. При этом в ручьях на увалах выбирает участки, где течение слабое. Его неветвящиеся веточки коротки и поэтому, хотя он и образует заросли, они всегда выглядят прозрачными. Из всех хвощей он самый высокий, до 70 см.
В противоположность ему, камышковый - самый лесной, очень низкий, не более 15 см. Кроме того, стебли его извилистые, поэтому в траве малозаметные, напоминая клубочки, в которых только при внимательном рассмотрении видны темные спорангии.
Представители следующего класса, плауновые, в сравнении с папоротниками и хвощами, в зелени тайги не выделяются своими размерами, зато удивляют обликом. Они похожи на разветвленные ленты, которые стелются по самой земле, как бы плывут. В тайге обычны два вида: годичный и сплющенный.
Стебли годичного, или колючего, покрытые жесткими короткими листочками, походят на светло-зеленые мохнатые разветвленные ленты, расползающиеся от корня в стороны до метра. В июле на концах веточек поднимаются спорангии - желтоватые колоски, наполненные спорами. Созревшие споры в колосках обладают одной особенностью, на которую с древних времен обратил внимание человек - хорошо впитывать влагу и не слипаться, поэтому их использовали для заживления ран. Не потеряли своего значения споры и в наши дни: из них готовят детскую присыпку - ликоподиум.
Плаун сплюснутый еще менее заметен в зелени трав и мхов. Сплюснутые стебли покрыты прижатыми листочками, и только тонкие, поднимающиеся колоски делают растение более заметными.
Про плаун годичный следует сказать еще то, что на Севере его часто использовали в эстетических целях. Осенью, когда начиналась установка зимних внутренних рам в окна, его извилистые стебли закладывались вместе с кистями рябины между ними. Такое украшение, контрастируя с однообразной белизной снегов за окном, всю зиму радовало взор живыми красками лета.
Если представители папоротников, хвощей и плаунов, в большинстве случаев, в тайге встречаются островами или кружками, то представители класса мхов - сплошным ковром, стелющимся по земле. Мхов насчитывается более сотни видов, но люди различают между ними всего несколько видов, потому что они мелкие и выглядят одинаково, да и в хозяйстве мало применимы. Для того, чтобы увидеть различия между ними, надо низко наклониться или встать на колени, но не помогает и это: надо носить увеличительное стекло для рассматривания деталей строения. Из-за своих малых размеров и неброских различий редкие из мхов имеют запоминающиеся русские названия. Исключением из этого является один широко распространенный, с типичным обликом для этого семейства - кукушкин лен.
В систематическом плане он принадлежит к семейству политриховых, подкласса бриевых, для которых характерно преобладание видов, имеющих жесткие зеленые игольчатые листья.
С этим мхом, про который, как и про хвойный лес, можно сказать поговоркой - "зимой и летом одним цветом", мы встречаемся с весны до осени. Он становится особенно нужным в летнюю пору, когда, собирая ягоды, захочется отдохнуть и присесть на землю, здесь лучше сухой кочки, покрытой кукушкиным льном, нет. Делать это нужно как можно реже, при отсутствии пней и валежника, так как, несмотря на "выносливую" упругость, мох очень чувствителен к давлению, особенно частому, поэтому быстро исчезает на тропинках.
Людям кочки, покрытые этим мхом, всегда кажутся однообразно зелеными, на самом деле, это не так, стоит войти в лес в мае или начале июня, как издалека в них обнаружатся новые цвета: беловатые и коричневатые. С близкого расстояния будет видно, что коричневый цвет придают скопления тонких нитей, а беловатый - острые колпачки из серебристых волосков, торчащих на кончиках этих нитей. При взгляде на такую кочку, порой, в голову может прийти сравнение, что это какие-то отряды крошечных лесных существ, построившихся с поднятыми к небу древними пиками, приготовились к завоеванию новых земель. Такое фантастическое предположение о подготовке к завоеванию станет близким к реальности, если снять серебристые колпачки, под ними обнаружатся зеленые или коричневые, в зависимости от времени года, коробочки, набитые спорами, которые и будут завоевывать, вернее, заселять новые просторы тайги.
Сами коробочки, имеющие ребра и крышечку разнообразной формы, напоминающие высокую пивную кружку, имеют большое значение в различии разных видов мхов, наряду с другими деталями строения, однако большинство их можно рассмотреть только под лупой, а лучше в специальный прибор - бинокуляр.
Благодаря особенностям формы коробочек, хорошо запоминается еще один мох из этого же подкласса, правда, встречается он редко, так как для своей жизни выбирает особые условия - кучи помета крупных лесных животных. Здесь в пору спороношения он выглядит в виде красноватых зонтиков на таких же красноватых нитях, торчащих группами - это сплахнум красный. При первых встречах он может показаться представителем другого царства - грибов.
В отличие от представителей семейства политриховых мхов, виды семейства бриевых и родобиевых стебельки имеют короткие, поэтому напоминают многолучевые звездочки, разбросанные по сырой земле. Когда же начинается спороношение, они принимают особый вид, напоминая тропическую траву пампасов, у которой из пышного скопления листьев, расположенных кругом у земли высоко вздымаются цветочные побеги, порой до 1,5 метров. Чтобы сравняться с такой травой, наши мхи должны увеличиться в сотни раз.
Каждый человек, осматривая деревянные крыши домов, а теперь и шиферные, обращает внимание, что они со временем покрываются какими-то зелеными островками, вначале мелкими, затем более крупными - так проявляют себя мхи из рода туидьевых, с очень нежными и линейными листочками. Они так же, как другие мхи в период размножения, приобретают коричневую окраску, которую им придают крохотные коробочки на ниточках-ножках.
На крышах эти мхи получают простор и чувствуют себя выше других, при этом успешно разрастаются, однако для хозяев домов это не приносит радости. Перегной, накапливающийся под ними, задерживает влагу и ускоряет процесс гниения досок. Раньше для сохранения досок их пропитывали смолой, потом крыши стали крыть шифером, но мхи прижились и здесь.
В подклассе бриевых мхов, наряду с мелкими, в лесах встречаются более крупные и другой формы. Так, например, представитель семейства ретидиевых - птилий гребенчатый, растущий в основании древесных стволов, похож на желтоватое птичье перышко, достигает высоты 4-5 см.
Другой представитель рода хилокомиумов, предпочитающий сырые ельники, демонстрирует оригинальную форму своего строения, очень похожую на древесную сосновую с ветвями-мутовками, но очень малых размеров. Эти мутовки, как и у сосны, появляются каждый год, и мох получает очередной этаж зелени, поэтому его называют еще "этажный". Форму крошечных деревьев с развесистыми кронами демонстрирует еще один мох - клемациум, который предпочитает селиться на сухих полянах.
Вместе с деревоподобными мхами лес богат и другими формами, не имеющими "ствола", например, из рода туидиевых и леукодиевых. Они довольно часто встречаются в березниках, где, как рыхлая зеленая шерсть, собираются у основания стволов или разрастаются в нижних ветвях. Их крошечные коробочки со спорами так мелки, что даже в пору массового спороношения слабо меняют зеленый цвет.
Зеленые, или гипновые, мхи не только густо заселяют землю, но и воду, где господствуют представители семейств фонтиналисовых и каллиэргеновых групп "водников".
Фонтиналис нередко селится в постоянных ручьях чугаса, бегущих по оврагам, и летом, разрастаясь, проявляет себя зелеными нитями, которые при ближайшем рассмотрении предстают как нежные стебельки с листочками-волосками.
Коллиэргены, или "черные мхи", предпочитают спокойную воду и очень часто болотную. Именно здесь они создают опасные своей обманчивой зеленью "полянки", которые скрывают глубокие ямы. Этому способствует то, что подводное переплетение стеблей с черными листьями походит на торфяной грунт, а верхние зеленые кончики - на траву, растущую из него; не замечая этого, человек или животное подходят к болоту, смело вступают на такую "полянку" и вдруг оказываются в черной грязи, которая уходит из-под ног.
Тесно с влагой связана еще одна группа мхов из подкласса бриевых, получивших собирательное русское название "серебрянок". Такое название возникло из-за их особенности, с одной стороны, когда листочки погружены в воду и удерживают на себе пузырьки воздуха, то под ее поверхностью они приобретают серебристый вид. С другой стороны, на суше, где они обычно растут рядом со стволами на сырых местах, подолгу сохраняют капельки росы или дождя, тоже серебристого цвета. Листья этих мхов плоские и походят на листья обычных трав, но имеют несколько особенностей. Первая - они очень мелкие, от нескольких миллиметров до одного, и тонкие, меньше миллиметра. При такой толщине в них с помощью микроскопа можно изучить строение и жизнь одной клетки. Еще одна характерная особенность жилок, которые, как сеть, покрывают листья цветковых растений, у них остаются по одной, а сами листочки имеют обыкновение равномерно уменьшаться на веточке от основания к вершине.
Если представители подкласса юнгерманиевых мхов практически выпадают из поля зрения собирателя лесных даров из-за своих мелких размеров, а также сходства с другими, то представители подкласса маршанциевых могут привлечь внимание своей формой при более крупных размерах. Из них довольно обычна по берегам ручьев прессия. Ее плоские разветвленные тела, извиваясь по самой земле, с весны украшаются очень своеобразными спорангиями, похожими на звездчатые зонтики в диаметре до 6-7 мм и потому хорошо заметными.
В отличие от наземных форм маршанций, водные более мелкие, и форма их напоминает обрывки травы, плавающие среди другой зелени, практически не привлекает взгляда. К ним относится, например, ричия, длиной 4-5 мм, с разветвленными концами, которые походят на какой-то мусор.
Если для человека значение разнообразных гипновых мхов пока невелико, то для тайги - громадное. Именно они своим плотным ковром защищают почву от водной эрозии. Их покров предохраняет корни деревьев и кустов, расположенных близко у поверхности почвы, от низких температур, когда снежный покров не образовался или, наоборот, рано сошел. Они поселяются на сплошных гарях, создавая первые скопления гумуса. Они регулируют влажность и температуру почвы в летний период, не допуская резких колебаний. В тайге, где из-за плохой освещенности не возникает луговая дерновая почва с многолетним растительным войлоком, скрывающим дневную жизнь беспозвоночных и мелких позвоночных животных. Моховой покров берет на себя эту защитную роль. Наконец, зимой рыхлый незамерзающий моховый покров облегчает движение мелким мышевидным животным, не впадающим в спячку.
В хозяйстве человека гипновые мхи играли важную роль как утеплители жилища в период продвижения его на Север, особенно в древности. Сейчас эта роль осталась там, где идет деревянное строительство. Здесь сухие лесные мхи постоянно используются для утепления пазов, между бревен в оконных и дверных проемах.
В лесу, занимаясь поиском грибов или ягод, человек, перемещаясь с места на место, часто встречает острова мхов, как обычно мягких, но другого цвета, чаще бледно-желтого или буроватого, которые занимают различные понижения среди ровной поверхности грунтов.
Если присмотреться к этим желтым островам, то обнаружится, что растения, создающие их, походят на зеленые мхи. Стоит потянуть одно из них, как сразу обнаружится различие - они без труда отрываются, потому что не имеют корня. При близком рассмотрении вырванного стебелька обнаружится другая разница в общем строении. Их мелкие веточки с мелкими листочками, равномерно покрывающие стебель, наверху сгущаясь, образуют характерную звездчатую головку, которой нет у зеленых мхов. Такое строение характерно для сфагновых или торфяных мхов. Отсутствие корней у торфяных мхов накладывает большой отпечаток на их жизнь. Во-первых, воду они получают листьями, а не по стеблю от корней. Во-вторых, они, ежегодно нарастая верхними частями, как бы отбрасывают нижние, которые с годами, отмирая, превратятся в знакомые слои торфа.
Слой верхних живых мхов, называемых "очес", ежегодно подрастая и поднимаясь вверх, как дрожжевое тесто в кастрюльке, создает не виданные в других растительных сообществах экологические условия, связанные с угрозой быть погребенными с годами под торфом.
В округе встречается более десятка видов сфагновых мхов, и они внешне так похожи, что различить отдельные можно только под бинокуляром. В полевых условиях сделать это трудно, правда, некоторые виды выдает общая окраска - то красноватая, то зеленая, то желтая. Большинство видов живет на болотах, которые, благодаря сфагновым мхам, служат накопителями чистой воды и регуляторами водотока малых рек.
В лесах обычно встречается два вида: остролистный и компактный, остальные, например, магелланский, вульфа, бурый болотный, оттопыренный, характерны для болот.
Когда говорят о дремучести каких-то лесов, то обычно имеют в виду главный признак их - "седую замшелость", которую рождают растения, имеющие в облике мало общего с гипновыми и сфагновыми мхами, разве что малых размеров - это лишайники. Они представляют особый тип низших споровых растений, соединивших в себе типы водорослей и грибов в одном организме. Это соединение, получившее название симбиоз, дает им определенные преимущества перед другими.
В лишайниках гифы гриба, в первую очередь, создают ту или иную форму тела, его своеобразный "дом", в котором, поселяясь, зеленые водоросли придают ему зеленоватый цвет и, главное, накапливают энергию солнца. При таком сожительстве оба "хозяина" имеют выгоду: гифы гриба приносят минеральные вещества, а водоросли - органические. В конечном итоге лишайники приобретают особую способность жить в очень неблагоприятных условиях на Севере при длительных низких температурах. В результате только лишайники постоянно живут на голых, промерзающих зимой скалах Полярного Урала.
В тайге многие виды лишайников, которых в округе десятки, также живут в более жестких климатических условиях, а именно, на деревьях, подвергаясь зимой влиянию сильных холодов, летом - сухосте и жаре. Не смотря на это, процветают и создают картину дремучести и старости, которую подчеркивают данные им русские названия, бородачи и косматики.
Бородачи, или вислянки, принадлежат к нескольким родам, в лесу наиболее заметны своими бледно-зеленоватыми длинными телами. Отдельные виды их становятся рекордсменами среди всего мира лишайников. Один из них, уснея длиннейшая, достигает длины 1 метра. Кроме длины бородачи обладают еще одной редкой особенностью: они растут вниз к земле, а не вверх, как основная масса представителей зеленого царства.
Другая особенность бородачей - медленный рост, всего два-три миллиметра в год, присуща и другим лишайникам. За 60-70 лет, достигнув метровой длины, они практически проживают среднюю человеческую жизнь.
Третья особенность усней и бородачей также присуща большинству лишайников - высокие требования к чистоте окружающего воздуха. Стоит появиться в воздухе серному, а чаще углекислому газу, как они прекращают свой рост и гибнут, являясь, в данном случае, надежными указателями загрязнения воздуха.
В отличие от вислянок, лишайники, с общим русским названием косматки, растут в разных направлениях. Среди них эвернии, отличающиеся более широкими лопастями тела, тянутся преимущественно вверх и в стороны, тогда как более тонкие свисают вниз. Именно эти лишайники, разрастаясь вместе на сухих нижних ветвях деревьев, придают им неухоженный, "непричесанный", косматый вид.
Среди эпифитных, т. е. растущих на ветвях и стволах лишайников, как правило, присутствуют виды с плоскими и прижатыми телами - пармелии . Без их кружавчатых, голубоватых или зеленоватых тел лес бывает только при сильной загазованности. Там, где воздух чист, они, разрастаясь на темных стволах хвойных деревьев, особенно с северной стороны, придают всему лесу больше света. Кроме пармелий лопастную уплощенную форму демонстрируют и цетрарии, которые поселяются среди других лишайников, сразу выделяются своим золотистым цветом. Особенно ярким он бывает у редкой цетрарии, растущей на старых стволах можжевельника.
На древесных стволах, кроме лишайников с лопастными и кустистыми формами тела, есть обычные виды, состоящие из мелких бородавочек, чешуек, которые, соединяясь между собой, образуют хорошо видимые пятна и кружки, напоминающие накипь, за что их и зовут накипные.
Одним из таких накипных лишайников является стенная золотянка - характерный спутник осиновой коры. Если взять на себя труд и осмотреть стволы нескольких осин, то непременно встретишь на зеленой коре ярко- желтые пятна - это и будет золотянка. Часто рядом с ней встречается и другой накипной лишайник - колоплака, но черного цвета.
Благодаря своим малым размерам и, в большинстве, неяркой окраске, накипные лишайники дали много открытий новых видов, в том числе и на просторах округа. Так, в конце второго тысячелетия на территории Кондинского района были найдены эндемичные виды: гилекта Леушинская и гилекта чернеющая.
Разнообразию форм тела эпифетных лишаников не уступают и наземные. Именно среди них живет широко распространенный в округе лишайник, который в быту называют "мох", и который целиком поддерживает жизнь ценного для человека Севера копытного животного, а именно северного оленя, за что он и получил название кладония оленья.
Этот лишайник любит светлые сосновые леса, где, разрастаясь столетиями, создает сплошной, почти белый покров, из-за которого они получили название боры-беломошники. Видеть эти боры, особенно в летнюю жару - зрелище, запоминающееся надолго еще и потому, что их остается все меньше и меньше из-за вмешательства человека, который, в лучшем случае, вытаптывает их, в худшем - выжигает.
В жаркий день в таком лесу сразу поражает снежная белизна земли, из которой к небу тянутся желтые прямые стволы сосен, между которыми, в отличие от тайги, далеко просматривается все пространство. Тишина нарушается лишь какими-то шорохами, будто испуганные маленькие зверьки карабкаются по стволам, но это не белки, это шелестят всего лишь желтоватые тонкие пленки, свисающие с желтой коры под слабым ветром. Когда проходит первое восхищение от увиденной картины, и человек начинает идти дальше, его поразит новое явление - в жаркий полдень громкий хруст лишайников, как зимой хруст снега под ногами.
В тайге, где на земле господствуют влаголюбивые гипновые мхи, лишайникам трудно бывает найти сухие места, как в сосновых борах, поэтому здесь они не образуют больших зарослей и довольствуются старыми пнями, толстыми стволами поваленных деревьев или высокими сухими кочками, лишенными мхов.
По форме тела наземные лишайники делятся на две группы: кустистые и листоватые. Кладонии относятся к первой группе кустистых, среди них в тайге обычна лесная, а в борах - оленья. В лесу светло-зеленые, порой почти белые кустики кладоний своими веточками напоминают крошечные деревья, которые не сгибаются под ветром и, почти не меняя цвета в течение лета, выглядят неживыми, поэтому особенно неожиданной бывает встреча на кончиках веточек новых цветов. Наиболее яркими цветами среди кустистых лишайников раскрашиваются отдельные виды трубчатых кладоний, как, например, пальчиковый. У них в пору размножения самые кончики веточек вдруг приобретают ярко-красный цвет, как будто это капельки крови, упавшей из ран каких-то теплокровных животных.
В отличие от кустистых лишайников, листоватые предстают розетками или лентами. Среди листоватых наиболее крупная - лобария легочная, растущая на коре поваленных хвойных деревьев. Свое название, легочная, она, видимо, получила за сходство поверхности тела в широких жилах с внутренним строением легких млекопитающих.
Другие плоские лопастные лишайники, пельтигеры, которых несколько видов в тайге, встречаются чаще. Обычно они выставляют свои листовые тела возле пней или стволов деревьев, часто окружая их.
Значение лишайников в кругообороте энергии живого вещества особенно велико в горах, где они выступают в роли первых накопителей гумуса в трещинах скал, куда позже приходят цветковые растения. Ценны они как накопители гумуса и на равнинах, особенно в сосновых лесах, растущих на сухих песчаных грунтах
Высшие споровые растения
Работа по созданию участка "Высших споровых растений" в открытом грунте была начата в 1966 году по инициативе директора Ботанического сада проф. И.П. Белоконя. Современный коллекционный фонд собирали сотрудники нескольких поколений. Работы по созданию участка проводили сотрудники Е.Г. Ромс, Т.В. Плотникова, Г.Д. Ермоленко, первые растения из окрестностей г. Киева были привезены Т.П. Коршук. Курировала экспозицию и З.П. Козлова, которая была куратором коллекции архегониальных растений в закрытом грунте. Значительную научную и практическую работу с 1975 до 2000 года проводила к.б.н. Н.М. Стеценко, благодаря которой количество представленных в экспозиции растений увеличилось вдвое, и достигало 116 коллекционных единиц. На сегодняшний день куратором участка является к.б.н. Е.В. Вашека.
Пополнение коллекции происходило как за счет растений, привезенных из экспедиций, которые охватывали не только территорию Украины, но и районы Кавказа, Дальнего Востока, Средней Азии, так и за счет растений, полученных из других ботанических садов, и выращенных из спор. Последний способ в наше время является основным для увеличения коллекционных фондов.
По состоянию на конец 2006 года на участке "Высших споровых растений" представлено 125 коллекционных единиц, которые принадлежат к трем отделам (Lycopodiophyta, Equisetophyta, Polypodiophyta), четырем классам и четырнадцати семействам. Наиболее представлены папоротникообразные, которые насчитывают 84 вида, 1 подвид, 4 разновидности и 33 культивара.
Любители ботаники скептически улыбаются, когда слушают легенды о ночи на Ивана Купала. Как можно найти Кто это, вообще, придумал? Как можно найти то, чего никогда в природе не существовало?
Многие любители природы делят растения на цветковые и нецветковые виды. Нецветковыми называют споровые растения, примеры которых сегодня будут рассмотрены в статье.
Начнем знакомство с небольшого описания. Слово «спора» пришло к нам из греческого языка. В переводе оно означает «семя» или «семечка». Речь идет об очень маленьком образовании, размер которого - примерно 1 мкм.
Сформировались споровые растения очень давно. Фактически они - прямые потомки той флоры, которая попала из океана на сушу. К споровым растениям относится не только папоротник. Ученые подразделяют их на две категории: высшие и низшие. В первой категории - папоротники, плауны, мох и хвощи. Во второй - водоросли и лишайники.
Если говорить о высших споровых организмах, то у них очень интересный Здесь можно увидеть чередование особей бесполого и полового видов. Соответственно, и размножение, в зависимости от вида, идет половым или бесполым путем. Полный жизненный цикл непрерывен. Растение образовывает гаметофит (орган полового размножения) и спорофит (орган
Эволюция позволила развиваться этим видам растений в двух направлениях. Получились две обширные группы: гаплоидная и диплоидная. Описывая споровые растения, примеры гаплоидной группы которых - мхи, ученые доказали, что у них более развит половой гаметофит. Спорофит у гаплоидной группы имеет подчиненный статус. Диплоидное направление споровых организмов (хвощ и папоротники) имеет сильно развитый спорофит, а гаметофит в виде заростка.

Половое поколение всегда имеет антеридии и архегонии. Это мужские и женские органы. Мужские сперматозоиды подвижны, женская половая клетка статична. Чтобы ее оплодотворить, сперматозоид должен попасть в водную среду, по которой сможет добраться до цели. Оплодотворенная яйцеклетка образует зародыш, из которого прорастает неполовое поколение, то есть спорофит. Следующий этап размножения произойдет спорами, которые развиваются в спорангиях.
Не все знают, как отличить споровые растения. Примеры рассуждений на эту тему могут выглядеть так:
Если три признака присущи рассматриваемому растению, то это споровый вид.
Трудно найти человека, который никогда не видел папоротник. Это древнее растение используют для украшения парков и садовых участков. Комнатные виды папоротников выращивают в вазонах, а любители лесных прогулок много раз видели пышные и зеленые папоротниковые заросли.
У всех папоротников преобладают перисто-рассеченные Эти споровые растения (папоротники) имеют самые заметные спорангии. Место расположения этих органов - нижняя сторона листьев.
Для информации добавим, что папоротников в природе больше десяти тысяч. Все это разнообразие объединено в 300 родов.
Мхи можно отнести к наиболее примитивным видам высших растений. Все моховидные - это небольшие по размеру представители без проводящей ткани. Подразделение на стебель и листья у мхов условное. Эти удивительные споровые растения - примеры умения приспосабливаться к условиям природы.

Итак, тело мха условно разделено на стебель, листья и корни. Да, корни этому растению заменяют нитевидные выросты - ризоиды. Их основное отличие от настоящих корней - отсутствие У мха каждый ризоид - одна или несколько живых клеток.
Мхи хорошо чувствуют себя в болотистых местах, в тени или просто в сырости. Мхи активно испаряют влагу, но восполняют потерю всей поверхностью растения. Несмотря на то, что споровым растениям необходима вода, чтобы размножаться, некоторые способны пережидать периоды засухи, они даже приспособились выживать на скальных участках. Как это происходит, до конца не понятно.
Доминирующее поколение мхов - половое. Спорофит полностью зависим от гаметофита.
Споры не могут образовываться на листьях, как у папоротника, поскольку сами листья весьма условны. Для этих целей у мхов приспособлена споровая коробочка, возвышающаяся над половым гаметофитом на ножке-ниточке.
Особенностью мхов считается возможность вегетативного размножения. В этом процессе задействованы почки и клубеньки. Если вегетативную часть отделить от основного растения, то она развивается в самостоятельную особь.

Не будем перечислять все низшие споровые растения. Примеры, которые интересно описать, - водоросли. Эти растения не так многочисленны, как папоротники и лишайники, их чуть больше ста видов. Среда обитания этой флоры - вода. Водоросли не имеют листьев и корней. Крепятся к грунту или камням прозрачными крючками. Подразделяют водоросли на 11 отделов, 4 из которых человек научился использовать в своих целях.
Царство растения – Plantae, Vegetabilia
Общая характеристика царства
Представители царства – высокоспециализированные автотрофные организмы, питающиеся за счет процесса аэробного фотосинтеза. Их тело обычно расчленено на стебель, корень, лист, и они хорошо приспособлены к жизни в наземно-воздушной среде. Клетки растений имеют плотную клеточную стенку, в основе которой – целлюлоза. Основной запасной продукт – крахмал. Размножение вегетативное, бесполое (спорами) и половое (оогамия); мужские гаметы либо имеют ундулиподии (сперматозоиды), либо не имеют их (спермии). Характерно чередование полового (гаметофит) и бесполого поколения (спорофит) с преобладанием диплоидного бесполого поколения. Зигота у растений дает начало зародышу, который затем развивается в спорофит.
Царство растений включает не менее 300 тыс. видов (ныне существующих и вымерших), относящихся к 9 отделам – риниофиты (Rhyniophyta ) и зостерофиллофиты (Zosterophyllophyta ) (ныне вымершие), моховидные (Btyophyta), плауновидные (Lycopodiophyta ), псилотовидные (Psilotophyta ), хвощевидные (Equisetophyta ), папоротниковидные (Polypodiophyta ), голосеменные (Pinophyta) и покрытосеменные (Magnoliophyta ). Представители ныне существующих отделов, за исключением моховидных, характеризуются преобладанием в цикле развития бесполого поколения (спорофита), имеющего сосуды и (или) трахеиды. Благодаря последнему обстоятельству эти растения называют сосудистыми.
Растения делят на две группы: споровые и семенные . У споровых растений спорогенез и гаметогенез разобщены во времени и пространстве: спорофиты и гаметофиты являются отдельными физиологически самостоятельными организмами. Единицей размножения являются споры. У семенных растений гаметофиты сильно редуцированы и не являются физиологически самостоятельными организмами. Единицей размножения является семя.
Споровые растения являются первопоселенцами суши, которые дали начало в процессе эволюции семенным растениям.
Споровые растения
Включают следующие ныне существующие отделы: моховидные (Bryophyta ), плауновидные (Lycopodiophyta ), псилотовидные (Psilotophyta ), хвощевидные (Equisetophyta ), папоротниковидные (Polypodiophyta ).
Споровые растения появились в конце силурийского периода, более 400 млн. лет тому назад. Первые представители споровых были небольших размеров и имели простое строение, но уже у примитивных растений наблюдалась дифференциация на элементарные органы. Усовершенствованию органов соответствовало усложнение внутреннего строения и онтогенеза. В жизненном цикле происходит чередование полового и бесполого способов размножения и связанное с этим чередование поколений. Бесполое поколение представлено диплоидным спорофитом , половое – гаплоидным гаметофитом .
На спорофите образуются спорангии, внутри которых в результате мейотического деления образуются гаплоидные споры. Это мелкие, одноклеточные образования, лишенные жгутиков. Растения, у которых все споры одинаковые, называются равноспоровыми. У более высокоорганизованных групп споры двух типов: микроспоры (формируются в микроспорангиях), мегаспоры (формируются в мегаспорангиях). Это разноспоровые растения. При прорастании споры образуется гаметофит.
Полный жизненный цикл (от зиготы до зиготы) состоит из гаметофита (период от споры до зиготы) и спорофита (период от зиготы до образования спор). У плаунов, хвощей и папоротников эти фазы представляют собой как бы отдельные физиологически самостоятельные организмы. У мхов гаметофит является самостоятельной фазой жизненного цикла, а спорофит сведен до его своеобразного органа – спорогона (спорофит живет на гаметофите).
На гаметофите развиваются органы полового размножения: архегонии и антеридии . В архегониях , похожих на колбу, образуются яйцеклетки, а в мешковидных антеридиях - сперматозоиды. У равноспоровых растений гаметофиты обоеполые, у разноспоровых – однополые. Оплодотворение происходит лишь при наличии воды. При слиянии гамет образуется новая клетка – зигота с двойным набором хромосом (2n).
Отдел Моховидные – Bryophyta
Насчитывает до 27000 видов. Моховидные имеют тело либо в виде слоевища, либо расчленены на стебель и листья. Настоящих корней у них нет, их заменяют ризоиды. Проводящие ткани появляются лишь у высокоразвитых мхов. Частично обособлены ассимиляционная и механическая ткани.
В жизненном цикле доминирует гаметофит. Спорофит самостоятельно не существует, развивается и всегда находится на гаметофите, получая от него воду и питательные вещества. Спорофит представляет собой коробочку, где развивается спорангий, на ножке, связывающей ее с гаметофитом.
Размножаются мхи спорами, могут размножаться и вегетативным путем – отдельными участками тела или особыми выводковыми почками.
Отдел делится на три класса : Антоцеротовые (100 видов, шесть родов талломных растений), Печеночные и Листостебельные мхи.
Класс Печеночные мхи (Hepaticopsida )
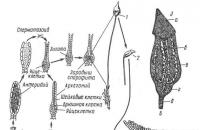
Класс насчитывает около 8500 видов. Это, в основном, слоевищные мхи, хотя есть виды, имеющие стебель и листья. Широко распространена маршанция обыкновенная (Marchantia polymorpha ) (рис 11. 1).
Рис. 11. 1. Цикл воспроизведения маршанции: 1 – таллом с мужскими подставками; 2 – таллом с женскими подставками; 3 – вертикальный разрез через мужскую подставку (в некоторых антеридиальных полостях находятся антеридии); 4 – антеридий в антеридиальной полости (н – ножка антеридия); 5 – двужгутиковый сперматозоид; 6 – вертикальный разрез через женскую подставку (а – архегоний).
Гаметофит имеет темно-зеленое слоевище (таллом), дихотомически разветвленное на широкие лопастные пластинки с дорсовентральной (спинно-брюшной) симметрией. Сверху и снизу слоевище покрыто эпидермой, внутри имеются ассимиляционная ткань и клетки, выполняющие проводящую и запасающую функции. К субстрату слоевище прикрепляется ризоидами . На верхней стороне слоевища в особых «корзиночках» образуются выводковые почки, служащие для вегетативного размножения.
Талломы раздельнополые, органы полового размножения развиваются на особых вертикальных ветвях-подставках.
Мужские гаметофиты имеют восьмилопастные подставки, на верхней стороне которых находятся антеридии . На женских гаметофитах подставки со звездчатыми дисками, на нижней стороне лучей звездочки расположены (шейкой вниз) архегонии. При наличии воды сперматозоиды перемещаются, попадают в архегоний и сливаются с яйцеклеткой.
После оплодотворения из зиготы развивается спорогон. Он имеет вид шаровидной коробочки на короткой ножке. Внутри коробочки, в результате мейоза, из спорогенной ткани, образуются споры. В благоприятных условиях споры прорастают, из них развивается протонема в виде небольшой нити, из верхушечной клетки которой развивается таллом маршанции.
Класс Листостебельные мхи (Bryopsida, или Musci ).
Листостебельные мхи распространены по всему земному шару, особенно в условиях холодного климата на сырых местах, в сосновых и еловых лесах, в тундре. На торфяных и моховых болотах часто образуют плотный ковер. Тело расчленено на стебель и листья, но настоящих корней нет, есть многоклеточные ризоиды. Класс состоит из трех подклассов: Бриевые, или Зеленые мхи; Сфагновые, или Белые мхи; Андреевые, или Черные мхи.
Андреевые мхи (три рода, 90 видов) распространены в холодных областях, внешне сходны с зелеными, по строению листьев и коробочки – со сфагновыми.
Подкласс Бриевые, или Зеленые мхи (Bryidae ). Насчитывает около 700 родов, объединяющих 14000 видов, широко распространенных повсюду, особенно в тундровой и лесной зонах Северного полушария.
Широко распространен кукушкин лен (Polytrichium commune ), образующий густые дерновины на сырых почвах в лесах, на болотах и лугах. Стебли до40 см высоты, неветвистые, с густыми жесткими и острыми листьями. От нижней части стебля отходят ризоиды.
Цикл развития кукушкина льна (рис. 11. 2).

Рис. 11. 2. Кукушкин лен: А – цикл развития мха; Б – коробочка: 1 – с колпачком, 2 – без колпачка, 3 – в разрезе (а –крышечка, б – урночка, в – спорангий, г – апофиза, д – ножка); В – поперечный разрез листа с ассимиляторами; Г – поперечный разрез стебля (ф – флоэма, крв – крахмалоносное влагалище, кор – кора, э – эпидерма, лс – листовые следы).
Гаметофиты кукушкина льна раздельнополые. Рано весной на верхушке мужских особей развиваются антеридии, на верхушках женских – архегонии.
Весной во время дождя или после росы сперматозоиды выходят из антеридия и проникают в архегоний, где сливаются с яйцеклеткой. Из зиготы здесь же, на верхушке женского гаметофита, вырастает спорофит (спорогон), имеющий вид коробочки на длинной ножке. Коробочка покрыта волосистым колпачком (калиптра) (остатком архегония). В коробочке – спорангий, где после мейоза образуются споры. Спора – небольшая клетка с двумя оболочками. На верхушке коробочки, по ее краю, расположены зубцы (перистом), которые, в зависимости от влажности воздуха, загибаются внутрь коробочки или отгибаются наружу, чем способствуют рассеиванию спор. Споры разносятся ветром и в благоприятных условиях прорастают, образуя протонему. Через некоторое время на протонеме образуются почки, из которых формируются листостебельные побеги. Эти побеги вместе с протонемой – гаплоидное поколение – гаметофит. Коробочка на ножке – диплоидное поколение – спорофит.
Подкласс Сфагновые, или Белые мхи (Sphagnidae)
К сфагновым мхам относятся свыше 300 видов единственного рода сфагнум (Sphagnum ) (рис. 11. 3).

Рис 11. 3. Сфагнум : 1 – внешний вид; 2 – верхушка ветви со спорогоном; 3 – спорогон (ш – остаток шейки архегония, кр – крышечка, сп – спорангий, кол – колонка, н – ножка спорогона, лн – ложная ножка); 4 – часть веточного листа (хлк – хлорофиллоносные клетки, вк – водоносные клетки, п – поры); 5 – поперечный разрез листа.
Ветвистые стебли сфагнума усеяны мелкими листьями. На верхушке главной оси боковые веточки образуют розетку почковидной формы. Особенностью сфагновых мхов является непрерывное нарастание стебля верхушкой и отмирание нижней части. Ризоиды отсутствуют, и поглощение воды с минеральными веществами происходит стеблями. Листья этих мхов состоят из двух типов клеток: 1) живых ассимилирующих, длинных и узких, хлорофиллоносных; 2) гиалиновых – мертвых, лишенных протопласта. Гиалиновые клетки легко заполняются водой и долго ее сохраняют. Благодаря такому строению, сфагновые мхи могут накапливать воды в 37 раз больше своей сухой массы. Разрастаясь плотными дернинами, сфагновые мхи способствуют заболачиванию почв. На болотах наслаивание отмерших частей мха приводит к образованию торфяников. Путем сухой перегонки из торфа получают воск, парафин, фенолы, аммиак; путем гидролиза – спирт. Торфяные плиты являются хорошим теплоизоляционным материалом. Сфагновые мхи обладают бактерицидными свойствами.
Отдел Плауновидные – Lycopodiophyta
Появление плауновидных связывают с силурийским периодом палеозойской эры. В настоящее время отдел представлен травянистыми растениями со стелющимися, дихотомически ветвящимися стеблями и корнями, а также спирально расположенными чешуйчатыми листьями. Листья возникли как выросты на стебле, и их называют микрофиллы . У плаунов есть флоэма, ксилема, перицикл.
Выделяют два современных класса: равноспоровые Плауновые и разноспоровые Полушниковые.
Класс Плауновые (Lycopodiopsida)
Из всего класса до настоящего времени сохранилось четыре рода.
Род плаун (Lycopodium). К этому роду относят многочисленные (около 200 видов) многолетние вечнозеленые травы, распространенные от арктических областей до тропиков. Так плаун булавовидный (L. clavatum) встречается в травяном покрове хвойных лесов на достаточно влажных, но бедных гумусом почвах. В сырых хвойных лесах широко распространен плаун годичный (L. annotinum ) (рис. 11. 4).

Рис. 11. 4. Плаун булавовидный.
Родбаранец (Huperzia). Представитель рода - баранец обыкновенный (H. selago) распространен в тундровой, лесотундровой и северной лесной зонах и произрастает в южно-таежных еловых лесах и ольшаниках, а также в мшистых лесах и на альпийских лугах.
Роддифазиаструм (Diphasiastrum ). Представитель рода дифазиаструм сплюснутый (D. complanatum) растет на сухих песчаных почвах в сосновых лесах.
Цикл развития на примере плауна булавовидного (рис. 11. 5).

Рис. 11. 5. Цикл развития плауна булавовидного :1 – спорофит; 2 – спорофилл со спорангием; 3 – спора; 4 – гаметофит с антеридиями и архегониями; 5 – развивающийся на гаметофите из зародыша молодой спорофит.
Стелющиеся побеги плауна булавовидного достигают до 25 см в высоту и более 3 м в длину. Стебли покрыты спирально расположенными ланцетно-линейными мелкими листьями. В конце лета на боковых побегах образуется обычно два спороносных колоска. Каждый колосок состоит из оси и мелких тонких спорофиллов – видоизмененных листьев, в основании которых расположены спорангии почковидной формы.
В спорангиях после редукционного деления клеток спорогенной ткани образуются одинаковой величины, одетые толстой желтой оболочкой гаплоидные споры. Они прорастают после периода покоя через 3-8 лет в обоеполые заростки, представляющие собой половое поколение и живущие сапротрофно в почве, в виде клубенька. От нижней поверхности отходят ризоиды. Через них в заросток врастают гифы гриба, образуя микоризу . В симбиозе с грибом, который обеспечивает питание, живет заросток, лишенный хлорофилла и неспособный к фотосинтезу. Заростки многолетние, развиваются очень медленно, лишь через 6-15 лет на них образуются архегонии и антеридии. Оплодотворение совершается при наличии воды. После оплодотворения яйцеклетки двужгутиковым сперматозоидом образуется зигота, которая без периода покоя прорастает в зародыш, развивающийся во взрослое растение.
В официнальной медицине споры плауновприменялись в качестве детской присыпки и обсыпки для пилюль. Побеги баранца обыкновенного используют для лечения больных, страдающих хроническим алкоголизмом.
Класс Полушниковые (Isoetopsida)
Селагинелла (Selaginella ) среди современных родов насчитывает самое большое (около 700) число видов.
Это нежное многолетнее травянистое растение, требующее высокой влажности. Селагинеллы в отличие от плаунов характеризуются разноспоровостью. В спороносных колосках образуется два вида спор – четыре мегаспоры в мегаспорангиях и многочисленные микроспоры в микроспорангиях. Из микроспоры образуется мужской гаметофит, состоящий из одной ризоидальной клетки и антеридия со сперматозоидами. Мегаспора развивается в женский гаметофит, не покидающий ее оболочки и состоящий из мелкоклеточной ткани, в которую погружены архегонии. После оплодотворения из яйцеклетки развивается зародыш, и затем новый спорофит.
Отдел Хвощевидные – Equisetophyta
Появились хвощи в верхнем девоне, достигли наибольшего разнообразия в карбоне, когда древесный ярус заболоченных тропических лесов в значительной степени состоял из древовидных хвощевидных, вымерших к началу мезозоя. Современные хвощевидные появились на Земле с мелового периода.
До настоящего времени сохранился лишь один род – хвощ (Equisetum), представленный 30-35 видами, распространенными на всех континентах.
У всех видов хвоща стебли имеют членистое строение с выраженным чередованием узлов и междоузлий. Листья редуцированы до чешуй и располагаются мутовками в узлах. Здесь же образуются и боковые ветви. Ассимилирующую функцию выполняют зеленые стебли, поверхность которых увеличивается ребристостью, стенки клеток эпидермы пропитаны кремнеземом. Подземная часть представлена сильно развитым корневищем, в узлах которого формируются придаточные корни. У хвоща полевого (Equisetum arvense) боковые ветви корневища служат местом отложения запасных веществ, а также органами вегетативного размножения (рис. 11. 6).

Рис. 11. 6. Хвощ полевой : а, б – вегетативный и спороносный побеги спорофита; в – спорангиофор со спорангиями; г, д – споры; е – мужской гаметофит с антеридиями; ж – сперматозоид; з – обоеполый гаметофит; и - архегонии.
Весной на обычных или специальных спороносных стеблях образуются колоски, состоящие из оси, которая несет особые структуры, имеющие вид шестигранных щитков (спорангиофоры ). Последние несут 6-8 спорангиев. Внутри спорангиев образуются споры, одетые толстой оболочкой, снабженной гигроскопичными лентовидными выростами – элатерами. Благодаря элатерам споры сцепляются вместе в комки, хлопья. Групповое распространение спор способствует тому, что при их прорастании разнополые заростки оказываются рядом, и это облегчает оплодотворение.
Заростки имеют вид небольшой длинно-лопастной зеленой пластинки с ризоидами на нижней поверхности. Мужские заростки меньше женских и несут по краям лопастей антеридии с многожгутиковыми сперматозоидами. На женских заростках в средней части развиваются архегонии. Оплодотворение происходит при наличии воды. Из зиготы развивается зародыш нового растения – спорофита.
В настоящее время большой роли в образовании растительного покрова хвощи не имеют. В лесах, на избыточно увлажненной почве, широко распространен хвощ лесной (E. sylvaticum) с сильно ветвящимися, поникающими боковыми веточками. На лугах, паровых полях, в посевах встречается трудно искореняемый сорняк хвощ полевой (E. arvense). У этого хвоща рано весной появляются неветвящиеся побеги, несущие спороносные колоски. Позднее от корневища развиваются зеленые вегетативные побеги. В лесной зоне на песчаных почвах и в оврагах широко распространен хвощ зимующий (E. hyemale ).
Вегетативные побеги хвоща полевого (E. arvense) в официнальной медицине применяют: в качестве мочегонного средства при отеках на почве сердечной недостаточности; при заболеваниях мочевого пузыря и мочевыводящих путей; как кровоостанавливающее средство при маточных кровотечениях; при некоторых формах туберкулеза.
Отдел Папоротниковидные – Polypodiophyta
Возникли папоротники в девоне, когда древовидные папоротники вместе с ныне ископаемыми плаунами и хвощами господствовали в растительном покрове земли. Большинство их вымерло, оставшиеся дали начало мезозойским формам, которые были представлены очень широко. Папоротники намного превосходят по числу современных видов все остальные отделы высших споровых (около 25000).
У большинства живущих сейчас (исключая тропические) папоротников отсутствует наземный прямостоячий стебель, но есть подземный в виде корневища. От корневища отходят придаточные корни и крупные листья (вайи ), имеющие стеблевое происхождение и длительно нарастающие верхушкой. Молодые листья обычно свернуты «улиткой». Среди существующих в настоящее время папоротников имеются как равноспоровые, так и разноспоровые.
В лесах Австралии, Южной Америки, Азии произрастают древовидные представители с колонновидными, неветвящимися, до 20 метров высоты стволами. В средней полосе нашей страны папоротники – это многолетние корневищные травы. Многие папоротники, как и мхи, являются индикаторами почв и типов леса. В светлых лесах, на песчаных или сухих подзолистых почвах распространен орляк обыкновенный (Pteridium aquilinum); на влажных богатых почвах кочедыжники (Athyrium) и крупныелесныещитовники (Dryopteris) (рис. 11. 7).

Рис. 11. 7. Щитовник мужской: А – спорофит: а – общий вид; б – сорусы на нижней стороне вайи; в – разрез соруса (1 – индузий, 2 – планцента, 3 – спорангий); г – спорангий (4 – кольцо); Б – гаметофит: 5 – сперматозоиды; 6 – заросток с нижней стороны (т – таллом, р – ризоиды, арх - архегонии, ан – антеридии); 7 – выход сперматозоидов из антеридия; 8 – архегоний с яйцеклеткой.
Цикл развития равноспоровых папоротников
В середине лета на нижней стороне зеленых листьев (у некоторых на специальных спороносных листьях) в виде коричневых бородавочек появляются группы спорангиев (сорусы) . Сорусы у многих папоротников прикрыты сверху своеобразным покрывалом - индузием. Спорангии образуются на особом выросте листа (плаценте) и имеют чечевицеобразную форму, длинные ножки и многоклеточные стенки. В спорангиях хорошо выражено механическое кольцо, которое имеет вид узкой несмыкающейся полосы, опоясывающей спорангий. Когда кольцо подсыхает, стенки спорангия разрываются, и споры высыпаются.
Споры, образовавшиеся в спорангиях, одноклеточны и имеют толстую оболочку. При созревании они разносятся током воздуха и в благоприятных условиях прорастают, образуя сердцевидную зеленую многоклеточную пластинку (заросток), прикрепляющуюся к почве ризоидами. Заросток представляет собой половое поколение папоротников (гаметофит). На нижней стороне заростка образуются антеридии (со сперматозоидами) и архегонии (с яйцеклетками). При наличии воды сперматозоиды проникают в архегонии и оплодотворяют яйцеклетки. Из зиготы развивается зародыш, имеющий все основные органы (корень, стебель, лист и особый орган – ножку, прикрепляющую его к заростку). Постепенно зародыш начинает существовать самостоятельно, а заросток отмирает.
У разноспоровых папоротников гаметофиты редуцированы до микроскопических размеров (особенно мужские).
Из корневищпапоротника мужского (Dryopteris filix-mas), получают густой экстракт, который является эффективным противоглистным средством (ленточные черви).

Предприятия уплачивают авансовые платежи по налогу на прибыль ежемесячно либо ежеквартально. Периодичность уплаты налога...
Включают следующие ныне существующие отделы: моховидные (Bryophyta ), плауновидные (Lycopodiophyta ), псилотовидные...

Татуировка не просто украшает тело – в ней заключен глубокий смысл, который человек стремится донести до окружающих. Тату...

Под строительство дома, можно начинать работы не дожидаясь, пока ребенку будет три года : для выплаты семье в трехлетие...

Рыба на гриле - невероятно вкусное блюдо. В нем соединяется аппетитная хрустящая корочка и нежная мякоть. Грилирование...

Мне так лень готовить! Я не думала, что когда-то такое напишу, но факты - вещь упорная. Мне лень. Готовить лень, а вкусно...

Цельные тыквенные плоды запекают в духовом шкафу, пользуясь их замечательно удобной формой. Надо же так было...

Если вы никогда не пробовали грузинский соус ткемали , вы многое потеряли. На самом деле приготовить этот...

Умерший человек, с момента смерти которого прошло не более 40 дней, считается новопреставленным. Считается, что...

В канун Рождества начинается лучшее время для гаданий, оно продлится до самого Крещения (19 января). Считается,...

Сами же последователи Никона, пользуясь государственной властью и силой, провозгласили свою церковь...

Ингредиенты курица и грибы отлично сочетаются, поэтому из них выходят вкусные блюда. Есть разные способы...

Совместимость мужчина Змея и женщина Свинья может сложиться только на основе общих интересов. Их отношения,...

Томатный сок - кладезь витаминов А и С, а также калия и других благоприятных для работы сердечно-сосудистой...
Включают следующие ныне существующие отделы: моховидные (Bryophyta ), плауновидные (Lycopodiophyta ),...

Татуировка не просто украшает тело – в ней заключен глубокий смысл, который человек стремится донести до...